If one amoeba divides into two, and two divide into four, and so on; how come we are not up to our ears in amoebas?
It is an established fact that cells reproduce by dividing from one cell into two. This does not seem to be a very profound statement to be making in the atomic age, but it is the key to what follows. If one cell divides into two, and the two resulting cells divide into four, and the four resulting cells divide into eight, we have what is known as exponential (or logarithmic) growth. This form of growth by doubling can have some profound effects. I have drawn a little chart that shows what will happen if a single cell continues to double at the rate of one every 24 hours.
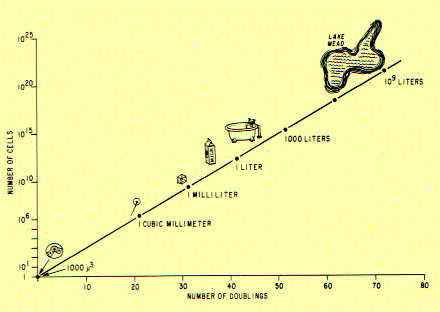
As you can see, nothing discernible happens for a while; but by 20 days, the single cell has grown to a mass the size of the head of a pin; by 30 days, it's the size of a small die (half of a pair of dice) --and after that, the mass of cells appears to explode, so that 40 days after the start, it is a liter, ten days after that it's a thousand liters (264 gallons) ,and by about 120 days (from a single cell) it's approximately the size of the earth.
This "explosion" in the size of a tumor (which is the simple consequence of a constant rate of cell division, and the fact that one cell divides into two) has led to the misconception that tumor cells divide faster as they get older. While this occasionally happens (tumor cells actually do start to divide faster), more often than not this "explosion" in size is simply due to the exponential way that tumors grow. This exponential growth pattern has also led to a number of other misconceptions, such as the idea that once a tumor is cut into, it grows faster. The cutting into it is probably coincidental, and if the tumor were not cut into, it would probably grow in the same way. In other words, most of this so-called tumor explosion can be accounted for by exponential growth --a consequence of the simple fact that one cell divides into two.
Early embryos grow exponentially, as do their organs. Obviously, this exponential growth doesn't last for very long. If an individual continued to grow at the same rate as he grew as an embryo the whole world could not contain him. What initiates the division of cells, and what stops them, is one of the great mysteries in biology. The answer is not known. There have been some interesting observations about what conditions are necessary to start and stop growth, but we still know little about the actual mechanisms.
We know, for example, that the potential for growth exists in most cells of the body. If you cut out a small piece of skin, the cells surrounding it divide and close up the hole. The same thing happens with connective tissue. If you take out a part of the liver, the rest of the liver will grow to achieve almost the same size as the original organ. The same thing is true of the adrenal gland and a number of other tissues. However, nerve cells apparently do not reproduce in the adult, nor do muscle cells (cancers of nerve and muscle cells are uncommon).
Michael Abercrombie found that cells grown in a culture dish will stop dividing when they form a layer that covers the bottom of the dish. If you cut out a wedge of cells, the adjacent cells will start dividing again to fill up the gap. We don't understand how the cells manage to know what to do.
There are feedback mechanisms which control the production of certain cells and keep the numbers of cells constant. For example, the loss of red blood cells due to bleeding or disease results in the kidney secreting a hormone called erithropoietin, which stimulates the production of more red blood cells. There are other mechanisms being investigated which are concerned with regulating the numbers of white blood cells in the blood.
Our bodies must respond to a large number of different conditions. The skin has to respond to temperature, the adrenal to various stresses, the ovaries have to undergo cyclic change, the child has to grow up to a certain age and then stop, growing bones have to grow to a certain length after which they have to harden and develop in such a way as to provide the best possible resistance to physical stresses, and so on. The timetables for initiating these changes are all different. Even if we should find that the cell has a single mechanism for responding to demands for growth and reproduction, we can be reasonably sure that whatever triggers the response will be different for different cells and organs. What is more, there is ample evidence now that we can expect wide differences in response from individual to individual. An interesting example of this individuality can be found if we observe what happens to different strains of mice when their ovaries are removed; a condition which simulates what happens when the ovaries have their function reduced by old age.
There are different strains of mice which are so highly inbred that they are similar to identical twins. What inbreeding in mice has done is essentially to take one genetic constitution and multiply it into a strain. Members of the same strain can exchange skin and organ grafts without reacting against each other. The rates at which they develop specific kinds of tumors is also relatively constant.
Let us use female mice from three inbred strains; we can call them strains A,B,C. (For aficionados of inbred mice, strain "A" represents C57BL, strain "B" is the C3H strain, and "C" is the CE strain.) If we take the ovaries out of mice of strain "A" we will find that all of the signs of sex hormone activity soon disappear. The uterus shrinks in size and the cells of the vagina show the distinctive signs of the absence of all of the sex hormones produced by the ovary. If we do the same thing with strain "B" we get a slight reduction in size of the uterus, and the cells of the vagina change to indicate the absence of hormones; but by three months the uterus is back to normal size and the cells of the vagina show ample evidence of hormone stimulation. Some even show signs of the typical cyclic changes that characterize the mouse's sexual cycle. It can be shown that the adrenal gland is producing the hormones that we usually associate with the ovary and that it is doing this in response to the pituitary hormones that ordinarily also stimulate the ovary (the gonadotropins). Strain "C" shows an even more fascinating response. It undergoes the same general changes as in strain "B"; only this time we find that not only does the adrenal cortex (the outer part of the gland) change its appearance and start producing sex hormones, but almost every animal will develop a cancer of the adrenal cortex within a year after the operation. Robert Huseby found that the differences between these strains is not a difference in the hormones produced, nor in the animal in general; but it is a very specific difference in the cells of the adrenal cortex itself.
It is reasonable to suspect that these kinds of differences between individuals and between strains of animals are characteristic of all living things, and probably occur in every organ in an animal. The discovery of a regulatory mechanism that appears to be ubiquitous is generally followed by someone finding an exception to the rule. Cancers themselves are highly individual. Cancers that have arisen in individual mice of one strain should all be genetically identical. Yet, each cancer behaves quite differently, even though they look alike and are produced by the same agent. One tumor may grow very slowly, and another very rapidly. One tumor may remain in the same place, and the other metastasize very rapidly. A tumor can arise in an animal, and be transplanted from one animal to another animal of the same strain. In the course of transplantation it may suddenly start to grow at a more rapid rate and it continues to grow at the accelerated rate.
We know that the growth of normal breast tissue can be accelerated or retarded by sex hormones. The same applies to tumors, except that this matter of individuality comes into play. This is true in tumors of both mice and people. The removal of the ovaries will slow down one tumor, and do nothing to another; the injection of hormones may accelerate one tumor and do nothing to another. One might say that breast tumors have personalities and respond in very different ways to different stimuli. What is the reason for this highly variable response? No one knows.
My first contact with the word cancer was a description which described cancer as consisting of "wildly growing cells." Compared to bacteria or embryos, they're pretty tame. Cancer cells often reproduce at much slower rates than the tissue from which they arose. Most tumors of the intestine have much slower cell division rates than the normal intestine. The "wildness" of tumors really relates, as does the wildness of teenagers, to their not knowing their place. The tumors may no longer respond to the influences that tell them either when to stop growing or in which direction to grow, or to stay in one place and not move all over the body where they don't belong. In the same way, some breast tumor cells are very responsive to sex hormones, or the lack of them, and some pay no mind at all.
There is, of course, a superficial similarity in the ways that everything responds, and it is a matter of taste whether you consider the similarities to be more important than the differences.In the past many scientists have equated cell division with tumor growth. There are some tumor systems where this is true, but there are also many where it is not. In tumors of the intestine, the rate of cell division is generally slower than the rate of cell division in the normal intestine. The reason that these tumors are malignant is because the cells invade the "inside" of the animal instead of obligingly sluffing off into the lumen of the intestine (the word "lumen" is a way of referring to the empty space surrounded by the intestine, which enables us to avoid calling it the hole inside. The part of the hose through which the water goes is the lumen). In the case of cancer of the intestine, therefore, one of the major problems is that cell loss does not occur as it occurs in the normal intestine. This, coupled with the fact that the tumor cells refuse to remain in one place, makes these tumors pretty serious, even though the cell division rate is slower than the rate in a normal intestine.
Apparently, most tissues have the capacity to grow rapidly, The growth of connective tissue around bone following a fracture resembles a malignant bone tumor enough so that it could confuse a pathologist into diagnosing it as cancer if he didn't know that there was a fracture present. Fortunately, physicians don't generally send pieces of regenerating bone to pathologists. The rate at which tissue grows around a fracture is phenomenal. The same thing is true of injured skin or intestine. The ability to reproduce is inherent in all living cells, The mystery is why they don't continue to grow. If we could understand the mechanism that stops growth, maybe the growth of some tumors might be controlled. Since we really don't understand what makes normal skin cells divide when they've been injured, nor what makes connective tissue cells divide when they're injured, nor what makes bone regenerate, and a whole wide variety of normal processes; you can see that we're very poorly equipped to cope with the changes that occur in cancer.